Zhanna Reznikova*, Boris Ryabko**
* Novosibirsk State University and
Institute for Animal Systematics and Ecology
Siberian Branch RAS, Frunze str. 11, Novosibirsk, 630091, Russia
e-mail: ryabko@neic.nsk.su
tel. 7 3832 284938
** Siberian State University of Telecommunication and Computer Science
Kirov Str. 86, Novosibirsk, 630102, Russia
The main point of proposed approach to study ants’ cognitive abilities is that our experiments provide a situation in which insects have to transmit information quantitatively known to the experimentalist in order to obtain food. One may estimate some properties of ant intelligence by measuring complexity of tasks they solve in order to pass definite pieces of information from scouts to foragers.
Our previous experiments, basing on ideas of Information Theory, have
shown that ants are able to memorize and transmit messages concerning sequence
of turns toward a trough of syrup and use the simplest regularities to
compress the information. To reveal counting and number related skills,
we suggested red wood ants Formica polyctena to transmit information
on the number and coordinates of objects. One of the experimental set-ups
consisted of a “tree trunk” with branches that ended in empty troughs,
except for one which was filled with syrup. Another set-up consisted of
a lattice which simulated Cartesian coordinates. The foragers of F.
polyctena were separated into teams of 5-8 individuals , each with
one scout. All laboratory ants were marked with coloured labels. To start
the experiment, an ant scout was placed at the randomly numbered trough
containing food and then returned to the nest on its own. The duration
of the contact between foragers and the scout was measured. Then we removed
the scout and the foragers had to search for the food by themselves. The
experiments were so devised as to eliminate all possible ways that may
help to find food, except for distant homing. It turns out that the ants
are able to count within several tens, and transmit this information to
their nestmates. The analysis of time duration of ants’ contacts enable
us to create a hypothesis of how they use numbers and coordinates in their
communication. We suppose that only a few highly social ant species use
such a complex communication system based on cognitive processes. At the
same time, we believe that the experimental schemes described can be used
to study the communication systems and numerical competence of other animals.
The communication systems of animals have been a matter of special
research interest to ethologists. Studying communicative means of different
species is a good tool to judge about their cognitive abilities. Direct
dialog with apes and grey parrots based on adopted human languages has
revealed astonishing mental skills such as ability to abstract, to extrapolate,
to count, to plan behaviour, and to use the experience gained in new situations
(Gardner, Gardner,1969; Pepperberg, 1987; Savage-Rumbaugh and Lewin , 1994
and others). At the same time, surprisingly few is known yet about natural
communication systems in animals. Although it is intuitively clear that
many high social species have to possess complex language, only two types
of natural communications have been decoded up to the present. First, one
of the most complicated of the known natural “languages ” in animals is
the symbolic honey bee “ Dance language”. Based on a distant homing system,
discovered by K. Von Frisch ( 1923, 1950) it was later intensively studied
using different methods including robotics (Michelsen, 1993 ). The second
communicative system being deciphered concerned acoustic signals of danger
which were deciphered for vervet monkeys (Struhsaker , 1967; Seyfarht et
al., 1980 ) and later for several other species ( Pereira , Macedonia ,
1991; Slobodchikoff et al., 1991 and others).
The main difficulties in the analysis of animal “languages” appear to be methodological. Many workers have tried to directly decipher animal language by looking for “letters” and “words” and by compiling “dictionaries”. The fact that investigators have managed to compile such “dictionaries” for a few species only, appears to indicate not that other animals lack “languages”, but that adequate methods are lacking.
As early as at the end of the 40’s, C.E. Shannon ( 1948) developed the basis of the information theory. The fundamental role of this theory was appreciated immediately not only in the development of the technology of the information transmission , but also in the study of “natural” communication systems. It is intrinsic needs of using information theory in investigations of language behaviour, because this theory presents general principles and methods for developing effective and reliable communication systems. In particular, in the 50’s and 60’s the entropies ( degree of uncertainty and diversity) of most European languages were estimated. Information theory was used to estimate the quantitative capacity of the honeybee’s memory to memorize the location of a food source ( Wilson, 1971). But this has not been extended for another species and situations.
We have suggested a new approach to study quantitative characteristics of communicative systems and important properties of animal intelligence. The main point of this approach is not to decipher signals but to investigate just the process of information transmission by measuring time duration which the animals spend on transmitting messages of definite lengths and complexities. This allows to estimate intellectual potentials by observing the communicative process. Our experiments based on ideas of Information Theory have shown that ants probably have an even more intricate form of communication than the honeybee. We also succeeded in studying some properties of insect cognitive capacities , namely their ability to perform limited counting and to memorize simple regularities, thus compressing the information available ( Reznikova, Ryabko, 1993, 1994, 2000; Ryabko, Reznikova, 1996).
In this paper we describe experiments which demonstrate red wood ants as being able to count within several tens, and transmit this information to their nestmates. The analysis of time duration of ants’ contacts enable us to create a hypothesis of how they use numbers and coordinates in their communication. We suppose that only a few highly social ant species use such a complex communication system based on cognitive processes. At the same time, we believe that the experimental schemes described can be used to study the communication systems and numerical competence of other species of animals.
The experiments were performed on Formica polyctena as a
group retrieving species with a high level of social organization. In various
years 5 colonies of this species were used. Ants lived in a laboratory
arena (200 x 200 cm) , in a transparent nest ( 10 x 20 cm ) . The arena
was divided into two sections: a smaller one, containing a laboratory nest,
and a bigger one with an experimental system ( see details in: Reznikova,
Ryabko, 1994). Both sections were connected by a plastic bridge, that was
from time to time removed to modify the setup or isolate the ants . The
colonies consisted of about 2000 ants. All experimental ants were marked
with colored labels and fed by sugar syrup only in the experimental setup
once every 2 - 3 days .
All series of laboratory experiments were performed by a common schedule:a situation was created where ants, to obtain food, had to transmit each other an amount of information known by the experimentalist. Therein the time spent by the scouting ant on contact with foragers was measured.
As this has been shown before ( Reznikova, Ryabko, 1994, 1996), the foragers of this species separated into “ teams ” of 5-8 specimens, each with one scout. As soon as the scout found food , it informed its foraging team. In each series of experiments we placed scouts on the place with the trough containing food, using tweezers. The scout returned to the nest on its own. Sometimes the scout contacted its team at once, and the group began moving towards the setup. In this case, after the scout contacted the foragers, we removed the scout and the foragers had to search for the food by themselves. But more often, after the scout returned to the nest, it and returned to the trough alone. Sometimes it made errors and only found the food-containing trough after visiting some empty ones. Then it returned to the nest again, contacted only one team, and either remained with the team or left it. In the first case the scout was isolated and we watched its foraging team. In the latter case we let the scout to repeat its trips, sometimes up to four times, before it was able to mobilize the foragers.
In all the cases of mobilization , we measured the duration ( in seconds) of the contact between the scout and the foragers together in the transparent laboratory nest. We considered the beginning of contact to occur when the scout touched the first forager ant, while we took the end of contact to be the moment when the nest was abandoned by the first two foragers . Contacts were followed by numerous antennal movements. Scouts touched one to four foragers in turn, sometimes two simultaneously. When the scout repeatedly returned to the trough alone, we measured the duration of each of its contacts with foragers. Only the duration of the last contact, followed by the foragers' abandonment of the nest, was taken into account. As a rule, all of the previous contacts were brief ( 1-5 sec.) and resulted in food exchanges.
Each series of experiments was preceded by “training ” stages as long as 4 weeks, during which the ants explored setups and got accustomed to the work regime. At that time, we picked out actively working ants and marked them.
Then, “incapable” scouts which either could not memorize well the way to the trough or could not mobilize their groups were revealed and discarded from further experiments.
The experiments were so devised as to eliminate all possible ways helpful to finding the food , except distant homing, i.e. information contact with a scout. During contact between a scout and foragers in their nest, the experimental apparatus was replaced by a similar one with all troughs empty to avoid the use both of an odour track and the food odour. So, following their contact with the scout, the foragers visited troughs that were purpose fully empty. When a compact “team” reached a goal, they were given a trough with syrup immediately, i.e. the trough at the goal was filled. The search was considered a success when the team came to the correct place leaving behind not more than 1 ant. An unsuccessful search, when the team failed to come or came in a small number ( more than 1 forager behind) was called a failure.
The first series of 51 experiments conducted on a “comb-like” setup
consisted of a long horizontal plastic “ trunk” with 25 equally spaced
plain plastic branches, of 6 cm length, although horizontal ( fig. 1 ).
Each branch ended in an empty trough , except for one filled with syrup.
Ants came to the initial point of the trunk over an additional small bridge
( the setup was mounted on glass props covered with slippery oil to prevent
the ants from moving in a straight line ).
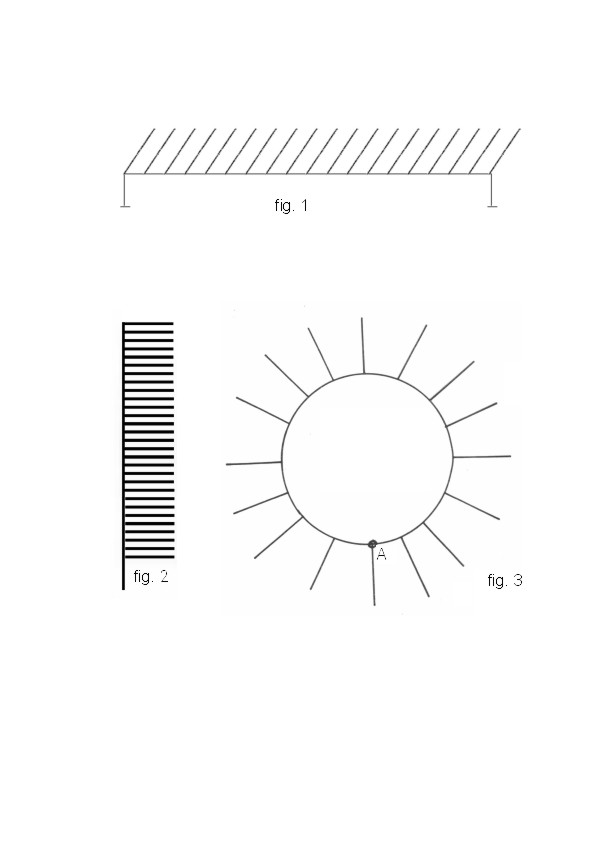
The second series of 31 experiments were carried out on a setup with vertically situated branches ( fig. 2 ). In order to test whether the time of transmission of information about the number of the “branch” depends on its length, as well as on the distance between the “branches”, another set of 16 experiments was carried out on a similar “vertical trunk” in which the distance between the “branches” was twice as great, and the “branches” themselves were three times and five times longer (for different series). The series of 38 experiments was conducted on a “circle-like” setup ( fig. 3 ).
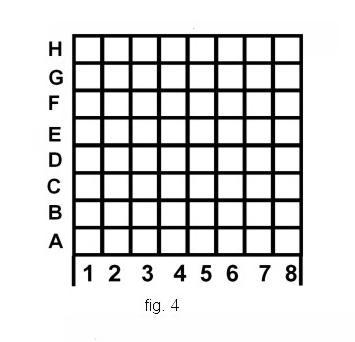
The investigation of the capacity of the ants for the transmission of information about coordinates of an object was carried out by means of a “Cartesian coordinate” setup, a flat grid of thin metal rods ( 8 by 8, 6 by 6 , 5 by 5 lines, fig. 4 ). Every setup was fixed so that the ants could only reach the starting point of the maze across a small bridge ( point A in fig. 3, 5 ). The further movements was possible only along the lattice rods. In one of the variants the lattice had a form of a “globe” with five “meridians” and five “parallels”; in this case the bridge led to the lower “pole” ( fig. 5 ). During the experiments the trough containing food was placed at various nodes of the lattice, while on the other nodes the empty troughs were placed.
In total, 32 scout-foragers groups ( “teams”) worked in three
kinds of setups. The teams abandoned the nests after they were contacted
with scouts and moved towards the trough lacking their guides 152 times
( note that we specially removed the scouts) . In 117 cases the team immediately
found the correct path to the trough, without making the wrong trips to
empty troughs. In the remaining cases, ants came to the empty troughs,
and began looking for food by checking neighbouring branches. In all experiments
( 31 in total) , foragers failed to find the food containing trough when
“ incapable” scouts were working. Such scouts were experimentally removed
from the working part of the arena.
Since all setups had no less than 25 branches, the probability to find the correct trough randomly is less than 1/25. Thus, the ratio we obtained experimentally can with high probability be accounted by information transmission from scouts. We note that the probability of finding the food-containing trough randomly in 117 cases out of 152 is less than 10 -10. In addition, ants , including scouts, placed in the setup, without having information on which trough contained food, usually failed to find the food, even though they actively searched for it.
|
|
containing branch |
forager contact |
Number |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Data obtained on the setup “vertical trunk” are shown in Table 1 as an example. In all setups the relation between the number of the branch i and the duration of the contact between the scout and the foragers t was linear, and may be described by the equality t = ai + b . The coefficient of correlation between t and i was high for different setups ( Table 2 ).
On the “circle” , the food-containing branch was numbered from the point
“A” counting clockwise, because in all cases the foragers moved only in
this direction, even when the counter-clockwise path was much shorter;
we do not know the reasons for this. It is also interesting to note that
on the “vertical trunk” after contact with the scout foragers climbed quickly
to the upper end of the trunk, and then slowly returned to the food - containing
branch; thus we numbered the branches starting from the upper end of the
trunk.
In total, 5 scout-foragers groups worked in 2 setups. The teams
abandoned the nests after they were contacted with scouts and moved towards
the trough 40 times ( note that we specially removed the scouts) . In 31
cases the team found the correct path to the trough in less than 5 min.
It is interesting to note that, contrary to situation when the ants had to choose the “right” branch, in a lattice the foragers moved from the initial point quickly, but not in a compact group, and reached the “node” with the trough each by its own pathway. Data on the time spent by the scouts on contact with their groups are presented in Table 3.
|
|
|
|
|
|
|
|
|
|
||||
|
“lattice” |
2 3 4 5 6 7 8 9 10 11 |
B6 B6 B6 D4 D4 D4 H4 H4 G2 G2 |
5 3 5 4 4 7 6 5 5 5 |
2 1 0 2 2 1 2 0 1 0 |
70 80 65 90 100 95 90 105 110 105 |
|
|
13 14 15 16 17 |
C2 C4 E2 E2 E2 |
7 5 6 5 6 |
0 0 0 1 1 |
55 70 44 52 48 |
|
|
19 20 21 22 |
C6 C6 C2 B1 |
4 5 6 4 |
0 1 2 0 |
45 40 50 56 |
|
|
24 25 26 27 28 29 30 31 |
B4 B4 D4 D4 D2 D2 D2 D2 |
4 4 4 4 6 5 3 3 |
1 0 0 0 1 2 0 0 |
72 65 50 58 45 52 50 50 |
|
|
|
|
|
|
|
8x8
“lattice” |
2 3 4 5 6 7 8 9 10 |
B6 B6 B6 D4 D4 D4 H4 H4 G2 |
7 10 6 12 4 1 9 5 8 |
- - - - - - - + - |
|
6x6
|
12 13 14 15 16 |
C2 C4 E2 E2 E2 |
8 6 4 9 2 |
- - - + - |
|
|
18 19 20 21 |
C6 C6 C2 B1 |
5 6 6 3 |
+ - - - |
|
|
23 24 25 26 27 28 29 30 |
B4 B4 D4 D4 D2 D2 D2 D2 |
9 10 11 10 6 7 12 3 |
+ - - - + - - - |
In order to test whether the foragers actually are guided by the information obtained from the scouts, a series of control experiments was carried out in which all of the ants were given the opportunity of access to an 8 by 8 “lattice”, on one of the “nodes” of which the trough with syrup was found, and were able to walk about it for 15 min ( let us recall that in the experiments the scout was purposely placed on the trough) . The results are presented in Table 4. A result in which during the allotted time ( 15 min.) at least one ant found the food, was considered positive. No one forager found food in 24 cases out of 30.
Let us introduce two statistical hypothesis. H0 : the probability of finding the food in which the foragers obtained the information from the scouts is the same as in the experiments with free search for the food; the alternative hypothesis H1 : the probability of finding food by “directed” foragers is greater than in cases of free search for the needed point. In order to test H0 against H1 we use the c2 test. Computations show that hypothesis H0 is rejected in favour of H1 at a significance level of 0.001. Thus, it follows from the data presented that ants are capable of transmitting the information about the coordinates of objects.
The most parsimonious explanation of results concerning ants ability
to search the “right” branch in the “comb”-like and “circle”- like setups
is that they can estimate the number of objects and transmit this information
to each other. Presumably they may pass messages not about the number of
the branch but about a distance to it or about the number of steps and
so on. Even if it is so, this shows ants as being able to use quantitative
values and pass the information about them. But it is worthy of noting
that the relation between the number of the branch and the duration of
the contact between the scout and the foragers is well described by the
equality t = ai + b for different setups which are characterized
by different shapes , distances between the branches and lengths of the
branches. The values of parameters a and b are close and
do not depend either on the lengths of the branches nor on other parameters.
All this enables us to suggest that ants transmit the information just
about the number of the branch.
The results of the experiments with “Cartesian coordinate” setup, showed that the ants are able to memorize and transmit information regarding the location of a trough containing food in a plane. Various mechanisms of coding or representation of such information can be hypothesized a priori: one hypothetical method is to number all the nodes of the lattice and to transmit the number of the node containing the food. Another method, which we have provisionally called the “coordinate” method is as follows: to memorize and transmit the number of the line and the number of the column at the intersection of which the food is found. Of course, other methods are possible as well.
Comparison of the results obtained with data regarding the time of transmission of information regarding the number of objects which was made above makes it possible to reject the first variant: in “numbering” all of the nodes of the lattice, the ants would have to transmit all the numbers from 1 to 64 (32 on average in the case of 8 by 8 setup). In order to transmit the number 32, 170-250 sec. would be spent by the scout, while in reality the time of contacts of the scout with the foragers was 60 - 150 sec. (87.2 on average), i.e. approximately two times less. In the cases of 6 by 6 and 5 by 5 lattices, the average time of transmission of information is also 2-2,5 times less than in the case of transmitting numbers 18 and 12 respectively. Thus, the data obtained make it possible to reject the hypothesis of the “numbering” of the cells of the lattice by the ants, and do not contradict the hypothesis of the use of “coordinate” method by them.
Summarizing, our data gained by applying the ideas of Information Theory, support the hypothesis that communicative means of red wood ants allow them to code and pass the information about numerical characteristics and coordinates of objects. This adjusts with recent data on coordination of working processes and division of labour within small working “teams” in this species in natural condition. It has revealed that, in order to obtain honeydew , a foraging group of 5 - 10 specimens has to search for a certain leaf with an aphid colony within such a huge 3- dimention space as a tree is for an ant. Successfulness of activity of these groups depends on activity of the scouts which attract “ shepherds”, “watch ants” and “transporting ants” to a new aphid colony ( Nongorodova, Reznikova, 1996; Reznikova, Novgorodova, 1998).
There are many situations in a history of studying of cognition in animals when applying an adequate method, animal behaviour students essentially changed imaginations about limits of intellectual possibilities of their subjects, particularly, of their numerical abilities. Thus, close treatment and examination of a chimpanzee ( Pan troglodytes ) applying a method of sample-choice, revealed its ability to count within limits of two objects only (Kohts, 1923 ). The use of sequential responding task, allowed to show a subject of the same species as being able to respond to arabic numerals between 1 and 9 and to suggest that an ape might form the integrated 9 - item linear representation (Matsuzawa, 1985; Tomonaga, Matsuzawa, 2000).
The numerical capacities of several avian and mammalian species have been examined since the beginning of the last century using a wide variety of numerical tasks. It has been shown that rats and birds, in particular, pigeons, crowns and parrots, are capable of executing a particular numbers of discrete responses, quite precisely when the required number of responses is low but not so precisely when the response requirement is high ( see Butterworth, 1999). Recently the use of symbolic numerosity stimuli allowed to demonstrate in pigeons a flexible constructive- counting -like behaviour that involved the production of up to six responses conditional upon the separate pre-presentation of up to six numerosity symbols ( Xia et. al, 2000). Crows turned to be able to choose the greater array from pairs of numerousness not only in the range of 1-12 but also 10 - 20 ( Zorina, 1997) although earlier they have been believed to discriminate objects within limits of 7 only ( Koeler, 1956). The use of animal’s natural “languages” for studying their intellectual capacities, particularly, numerical skills, amazingly has never been applied, although very impressed results have been achieved in this area by means of intermediate languages (Boysen, 1992; Pepperberg, 1994).
Studying processes of information transmission within ant’s colony may serve as a good background for starting with applying natural communicative means of animals to estimate limits of their cognitive abilities. As far as cognitive processes in social insects are concerned , excellent learning capacities have been demonstrated in ants and bees. Some experiments have shown these insects as being capable of abstraction, extrapolation and of solving rather complicated tasks in order to get bearings ( Reznikova, 1982; Collet et al.,1993; Lehrer, 1996). Recent studies have revealed a great deal of flexibility of foraging strategies in ants ( Mercier, Lenoir, 1999). At the same time, the bounds of insects’ numerical competence are still remain unclear. Only few findings concern this theme. Thus, at least, protocounting abilities of the honeybees have been demonstrated basing on experiments in which insects had to use number of landmarks as the criteria to find the feeder ( Chittka et al., 1995 ). It is also known that ants of several species are able to estimate quite precisely numbers of encounters with members of other colonies on their feeding territories ( Reznikova, 1980, 1994, 1999; Gordon, 1994; Brown, Gordon, 2000).
It is important to note that information transmission from the scouts to the foragers is possible only for a few high social ant species which possess structured cooperation and division of labour between leaders and followers. In spite of wide spread of solitary foraging in different ant species, collective behaviour and decision-making in the majority of species are based on numerous interactions among individuals which use local information and follow rather simple algorithms ( Detrain et al., 1999). Isolated instances have been described up to now concerning teams in the army ants Eciton burchelli ( Franks, 1996) and Dorylus wilverthi ( Franks et al., 1999) as well as teams in group retrieving Formica species including a key individual and followers ( Robson and Traniello, 1999). According to Anderson and Franks (in press) , “teams” revealed by ours (Reznikova, Ryabko, 1990, 1994, 1996), as well as “dance groups” in honey bees ( Waddington, 1989) should more properly be called “cliques”, sensu Holldobler and Wilson (1990:343) who define a clique as a “group of workers whose members recognize one another as individuals to accomplish some task”.
For ants, the bounds of applying of our approach are limited by set of species with social style of life basing on division of labour within “cliques”.
In conclusion, we believe that the elaborated method based on studying
of cognitive abilities, particularly, number related skills, by observing
the communication processes and by estimating the time necessary for information
transmission, can be extended for other social species, possessing flexible
behaviour, individual recognition and necessity to pass and memorize complex“messages”.
Supported by the Russian Foundation for Fundamental Investigations
( grants N 99- 01- 00586, 99-04-49713 and Russian Ministry of High Education
in the field of fundamental natural sciences е00-6.065.
We thank Prof. Donald Michie for fruitful discussion during his visit
to Novosibirsk in 1998.
Brown, M.J.F., Gordon ,D.M. ( 2000). How resources and encounters
affect the distribution of foraging activity in a seed-harvesting ants.
Behav.Ecol.Sociobiol.
,47, 195-203.
Boysen, S.T. (1992). Counting as the chimpanzee views it. In: Fetterman G., Honig, W.K. ( eds) Cognitive aspects of stimulus control. Erlbaum, Hillsdale, pp. 367- 383.
Butterworth ,B. (1999). The mathematical brain. Macmillan, London.
Chittka, L., Kunze ,J., Shipman, S., Buchmann, S. L. (1995) . The significance of Landmarks for Path Integration in Homing Honeybee Foragers. Naturwissenchaften, 82, 7, 342-343.
Collet, T. S , Fry, S.N., Wehner, R. (1993 ). Sequence learning by honeybees. J. Comp. Physiol., N. 172, pp. 693 - 706 .
Detrain, C., Deneubourg, J.-L., Pasteels, J.M. (1999). Decision-making in foraging by social insects. In: C. Detrain et al ( eds). Information Processing in Social Insects. Birkh? user Verlag Basel/ Switzerland. pp. 331-354.
Gardner, B.T. , Gardner, R.A. (1969). Teaching sign language to a chimpanzee. Science, N. 165, pp. 664 - 67 .
Franks, N. R. ( 1986) . Teams in social insects: group retrieval of prey by army ants (Eciton burchelli, Hymenoptera: Formicidae). Behav Ecol Sociobiol , 18, 425–429.
Franks, N. R., Sendova-Franks, A.B., Simmons, J., Mogie, M. (1999). Convergent evolution, superefficient teams and tempo in Old and New World army ants. Proc R Soc Lond B, 266, 1697–1701.
Frisch, K. Von ( 1923). ? ber die Spr? che der Bienen. Zool. Jahrb. 40, 1 - 119.
Frisch, K. Von (1950). Bees, their vision,chemical senses and language. Cornell Univ. Press, USA.
Holldobler , B. , Wilson, E.O. ( 1990). The Ants . Springer-Verlag, Berlin.
Koehler, O. (1956). Thinking without words. In: Proc. of the 14-th Internat. Congr. Of Zoology, Copenhagen, 1953, pp. 75 - 88.
Kohts, N. ( 1923). Utersuchungen ? ber die Erkenntnisf? higkeiten des Schimpanses, Museum Darwinianum, Moskau, p. 453.
Lehrer, M. (1996). Small - scale navigation in the Honeybee : active acquisition of visual information about the goal . The Journal of Exper. Biol., 199, 253 - 261.
Matsuzawa, T. (1985). Use of numbers by a chimpanzee, Nature, 315, 57-59.
Mercier , J.L., Lenoir, A. ( 1999). Individual flexibiloty and choice of foraging strategy in Polyrhachis laboriosa F. Smith ( Hymenoptera, Formicidae). Insectes soc., 46, 267 - 272.
Michelsen, A. (1993). The transfer of information in the dance language of honeybees: progress and problems. J. Comp. Physiol. A , Sensory, Neural and Behavioural Physiology., 173, 135-141.
Novgorodova ,T.A., Reznikova, Zh.I. 1996. Ecological aspects of interaction between ants and aphids in the forest park zone of the Novosibirsk Scientific Center. Siberian Journal of Ecology, 3-4, 239 - 245.
Pepperberg , I.M. (1987). Acquisition of the same/ different concept by an African Grey Parrot ( Psitacus eritacus ) : Learning with respect to categories of colour, shape and material. Animal Learning and Behavior, 15, Apr., 423 - 432.
Pepperberg, I.M. (1994). Numerical competence in an African grey parrot (Psyttacus erithacus). J. Comp. Psychol.. 108, 36 - 44.
Pereira, M.E., Macedonia, J.M. ( 1991). Ringtailed lemur antipredator calls denote predator class, not responce urgency. Animal Behaviour, 41, 543-544.
Reznikova , Zh. I. (1980). Interspecific hierarchy in steppe ants. Russian Journal of Zoology, 59, 8, 1168 - 1176.
Reznikova , Zh.I. ( 1982). Interspecific communication among ants. Behaviour (Leiden, Netherlands), 80, 1-2, 84-95.
Reznikova, Zh. ( 1994). The Original Pattern of Manegement in Ant Communities: Interspecies Social Control. In: Progress to meet the Challenge of Environmental Change. Manchester, UK, p.27.
Reznikova , Zh. I. (1999). Ethological mechanisms of population dynamic in species ant communities. Russian Journal of Ecology, 30, 3, 187-197.
Reznikova Zh.I., Novgorodova T.A. ( 1998). Division of labour and exchange of information within ant settlement. Advance in Modern Biology, RAS, 118, 3, 345 - 356 ( in Russian with English summary).
Reznikova, Zh. I., Ryabko , B.Ya. (1990). Information Theory approach to communication in ants. In: Sensory systems and communication in
Arthropods. Advances in Life Science, (K. Wiese et al. Eds) Birkh? user Verlag, Basel / Switzerland 305-307.
Reznikova, Zh.I., Ryabko, B. ( 1993) . Ants aptitude for the transmission of information on the number of objects. In: Sensory systems of Arthropods.
K.Wiese et al ( eds) Birkh? user Verlag Basel/ Switzerland. 634-639.
Reznikova, Zh. I., Ryabko, B. ( 1994) . Experimental study of the ants communication system with the application of the Information Theory approach. Memorabilia Zoologica, 48, 219-236.
Reznikova, Zh.I., Ryabko, B.Ya. ( 1996). Transmission of information regarding the quantitative characteristics of objects in ants. Neuroscience and Behavioural Physiology, 26, 5, 397 - 405.
Reznikova, Zh.I., Ryabko, B.Ya. ( 2000). Using Information Theory Approach to study the communication system and numerical competence in ants. In : From Animals to Animats 6. Proceeding of the Sixth International Conference on Simulation of Adaptive Behaviour. The MIT Press, Cambridge, Massachusetts, London, 501 - 506.
Robson, S.K., Traniello, J.F.A. ( 1999) . Key individuals and the organization of labor in ants. In: C. Detrain et al ( eds). Information Processing in Social Insects. Birkh? user Verlag Basel/ Switzerland. , 239 - 259.
Ryabko, B.Ya. , Reznikova, Zh. I. ( 1996) . Using Shannon Entropy and Kolmogorov Complexity to study the communicative system and cognitive capacities in ants. Complexity, John Wiley & Sons, Inc., New York, USA, 2, 2, 37- 42.
Savage-Rumbaugh, S. , Lewin, R. (1994). Kanzi: The Ape at the Brink of the Human Mind. New York: John Wiley & Sons.
Seyfarht, R.M., Cheney ,D.L , Marler, P. (1980). Vervet monkey alarm calls: semantic communication in a free-ranging primate. Animal Behaviour, 28, 1070 - 1094.
Shannon , C. (1948). A mathematical theory of communication. Bell System Techn. J., 27, 3, 379 - 443; 27, 4, 623 - 656 .
Slobodchikoff, C.N., Kiriazis, J., Fischer, C., Creff, E. (1991). Semantic information distinguishing individual predators in the alarm calls of Gunnison prairie dogs. Animal Behaviour, 42, pp. 713 - 719.
Struhsaker, T.T. (1967). Auditory communication among vervet monkeys ( Cecropithecus aethiops) . In: Social Communication Among Primates ( Ed. by S.A.Altmann), 281-324. Chicago: University of Chicago Press.
Tomonaga, M., Matsuzawa ,T. ( 2000). Sequential responding to arabic numerals with wold cards by the chimpanzee ( Pan troglodytes). Animal Cognition, 3, 1-11.
Xia , L., Siemann, M., Delius, J.D. (2000). Matching of numerical symbols with number of responses by pigeons. Animal Cognition, 3, 35 - 43.
Waddington, K. D. (1989). Implications of variation in worker body size for the honey bee recruitment system. J. Insect Behav ., 2, 91–103.
Wilson, E.O. (1971). The Insect Societies. Cambridge, MA: The Belknap Press, Harvard University Press, p. 548.
Zorina, Z.A. ( 1997). Reasoning in birds. Phys. Gen.Biol.Review, 11, 1-47.